
輕鬆簡讀版:
- 在自然的走、跑、跳之中,參與的肌肉不斷地再伸長與縮短,這類離心與向心收縮不斷接續進行的機制稱為伸展–收縮循環(SSC)。
- 單次跳躍屬於不連續動作,最常見的例子為下蹲跳(CMJ) {圖2},這類不連續的SSC運動始於離心階段的反向動作(countermovement) {圖1},之後才蓄力做出向心爆發;而連續動作則是會周而復始地應用到SSC,例如跑步和連續跳躍{圖9.}。
- 從離心轉換到向心收縮的重要性,這極短的轉換時間叫做攤還期(amortization phase){圖5.},攤還期太久或是之間有停頓,會導致離心階段儲存的彈性能量被肌肉和周圍組織吸收(部分轉成熱能)。
- SSC的前提是,作用肌的運動必須是等長(可有可無)->離心->向心收縮的順序,非這順序就不可能有SSC發生,例: 蹲踞跳(SJ)的動作順序為,離心->等長->向心收縮,因此並非是良好的SSC動作{圖4.}。
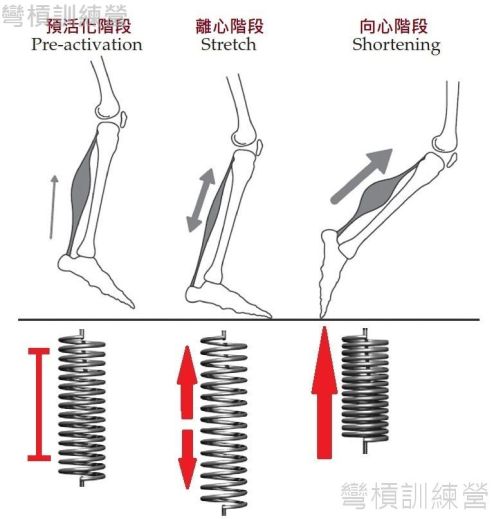
- 深跳(DJ) {圖3.}和跑步等等的連續動作都屬於完整的SSC運動,包含了三個重要階段,分別為: 預活化階段(pre-activation)、離心階段、向心階段{圖9.}。
- SSC對運動的效果就是使運動的向心發力階段的力量和速度提升{圖 7.}。詳情請看SSC的增益效果與力量–速度曲線部分。
- SSC的機制主要為1)動作在離心階段時,彈性組織儲存的彈性位能;2)離心階段讓肌肉作抗力,讓大量的橫橋能充分搭起;3)離心階段時對肌肉的牽拉導致牽張反射啟動,增加骨骼肌剛性,上述這些主要機制幫助操作者在向心階段時,能發揮出更強的的運動成果。 詳情請看伸展–收縮循環原理部分。
- 其它會應用到SSC的常見運動包括:跑步、田徑、舉重、足球等等運動,實際上是幾乎所有運動都有牽涉到SSC。
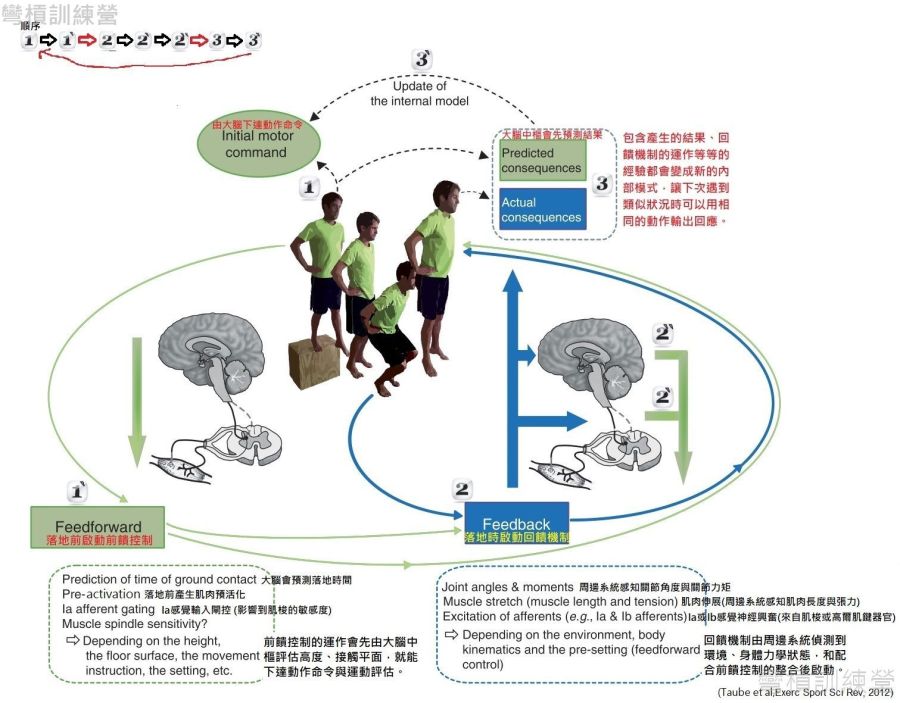
- SSC運動是前饋(feedfoward)控制與回饋(feedback)控制的整合運作{圖 20.}。回饋是運動過程中整合周邊反饋的信息並及時幫助身體調整當下動作與加固,如:組織儲存彈性張力、牽張反射;前饋控制代表人體在動作前所做的計畫或是預測,如: 運動前的評估、肌肉預活化。
文章中的常用文字縮寫:
伸展-收縮循環 (stretch-shortening cycle) : SSC
下蹲跳(countermovement jump): CMJ
深跳(depth jump): DJ
蹲踞跳(squat jump): SJ
肌電圖(electromyography): EMG
由於文章超過13000字,所以有目錄 (請讀者、粉蝨們善用ctrl+F,可以得到最棒的閱讀效果):
1. 正文與SSC的應用介紹
2. SSC的增益效果與力量-速度曲線
3. 伸展-收縮循環原理 (此部分有三個重要子題:
….*肌腱儲存的彈性位能
….*橫橋的活化態
….*神經的調控 )
4. 總結
5. 附錄 (此部分有三個重要子題:
….*牽張反射
….*梭內纖維會參與牽張反射提供給骨骼肌的增益性嗎?
….*預活化是前饋(feedforward)而反射是回饋(feedback)
6. 參考資料
正文與SSC的應用介紹:
伸展-收縮循環(Stretch-shortening cycle, 以下以SSC簡稱)在生活中是無所不在的,它使人能透過自主地或不自主地增進運動表現,本篇文章會介紹SSC的相關概念與應用方式。
對於活動量高的民眾,他們的生活中鮮少如拿起水杯喝水那樣簡單的事,因為日常生活中的活動都是由連續動作組成,走、跑等等讓人意想不到且平淡無奇的動作當屬最佳例子,而跳躍這種不連續動作也是經典案例。 在自然的走、跑、跳之中,參與的肌肉不斷地在伸長與縮短,這類離心與向心收縮不斷接續進行的機制稱為伸展-收縮循環(SSC)。
在不連續的動作中,SSC開始於反向運動(countermovement),於出力後結束 {圖1.}。這過程中,反向運動包含了主要作用肌的伸長(離心收縮),而在反向動作結束的當下,會有個為時極短的攤還期(amortization phase or immediate transition,轉換期)讓我們將離心收縮轉換到向心收縮,這短暫的轉換期事實上是一瞬間(幾百毫秒)的事情。 在反向運動的短暫時光裡,伸長的肌肉讓結締組織儲存了不少的能量,這些能量伴隨著向心收縮的力量一起展現,讓運動者能比單純做向心收縮還要發揮出更強的力量及更快的速度 [1]。
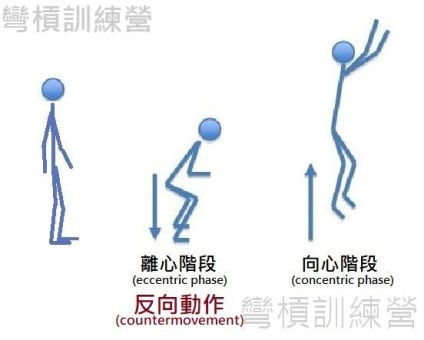
.
雖然觀眾都比較愛看運動員起飛的時候,但是離心階段是所有SSC運動的關鍵,有別於向心收縮是最後的結尾,離心收縮反而是動作的準備階段,離心收縮的好處除了在收縮過程中積蓄力量之外,他本身還是個有用的保護機制! 特別是在過程中包含了懸空到觸地的動作,離心收縮能透過伸長肌肉時一併抗力,避免觸地後重心(COG)的過度下沉。 離心收縮也幫助身體緩衝來自地面的衝擊,透過四肢、臀部、軀幹的肌肉一同合作,給出抵抗力量,讓力能沿著有剛性的身體平均分散,此舉能最大化地降低衝擊力由軟組織吸收的風險,可以有效地避免觸地時受傷。
.
以下提供兩個有應用到SSC的跳躍與一個沒用到的跳躍給大家參考 [2]:
*有應用到SSC的第一例是下蹲跳(countermovement jump, CMJ) {圖2},操作者再起跳前會先做出一個與跳躍方向相反的快速下蹲(countermovement squat),並馬上出力跳起,中間完全沒有刻意的停頓。

.
*另一例有用到SSC的是深跳(depth jump, DJ) {圖3.},深跳的一開始只是簡單的步下一定高度的椅子或箱子,並靠著操作者本身熟練的控制來做到輕輕落地,這下墜距離所產生的重力(很大的力量)必須靠著操作者做出一定幅度的下蹲來吸收,並在過程中沒停頓地馬上用力跳起; 與下蹲跳有些類似的地方是操作者落地時做出的下蹲正好是與跳躍方向相反的反向動作,只是深跳有抵抗更強的重力 [3]。

.
*而沒有應用到SSC的例子是蹲踞跳(squat jump, SJ) {圖4.},與下蹲跳不同的地方是操作者再起跳前,會下蹲到起跳位置並在該處停頓2~3秒後才出力起跳,這暫停時間到起跳前都必須維持身體的動作,而此停頓也讓下蹲變得不再是反向動作。

————————分隔線————————
.
眼尖的讀者應該已經知道後續要講什麼了,如果停頓時間太長就會影響到發力,深層的意義是這阻止了SSC的發生[2、3]。 相關機轉會在之後的段落解釋。 我們先來理解動作從離心轉換到向心收縮的重要性,這極短的轉換時間叫做攤還期(amortization phase,不同領域有不同稱呼)* {圖5.},這攤還期若是從高處落地或跑步時的踏地,那就會有極短暫的觸地時間(ground contact time),若動作是雙腳在起跳前都一直在地上的CMJ,那此動作的攤還期也常見用觸地時間或耦合時間(coupling time)做統稱。 攤還期太久或是期間有停頓(離心轉換到向心中間有等長運動),都很難有SSC產生。
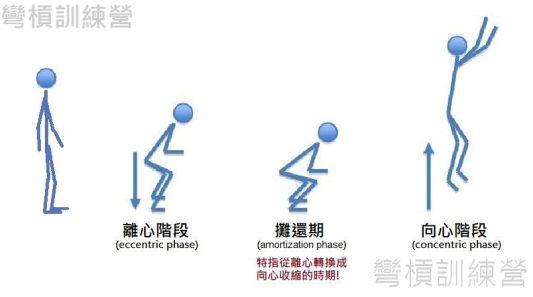
*[註1.]: 攤還期(amotization phase)指從離心動作到向心動作的階段,此階段因應各自的需求在不同領域有不同稱呼,例如:運動界除了稱之為攤還期外也稱之轉換期(immediate transition),也有神經科學家會以動作轉換的短暫停滯為稱呼(short delay),這動作轉換的時間也有部分科學家稱為耦合時間(coupling time)。本文章以攤還期做統稱 [1、2]。
.
由此可見,SSC有個一定的前提,就是作用肌的運動必須是等長(可有可無)->離心->向心收縮的順序,非這順序就不可能有SSC,例如:Depth Jump規定操作者下蹲後要停頓2~3秒鐘,那此時的肌肉動作順序為離心->等長->向心收縮,導致在離心階段儲存的彈性能量被肌肉和周圍組織吸收後(部分轉成熱能)就沒有增力效果了[4]。
以下表格附上部分常見運動的觸地時間或耦合時間,此表格為知名實務訓練家Owen Walker整理的表格{圖6.} [5]:
表格中清楚指出要引發強而有力的SSC就要有極短的觸地時間,這些常見的運動所呈現的觸地時間都相當短暫。
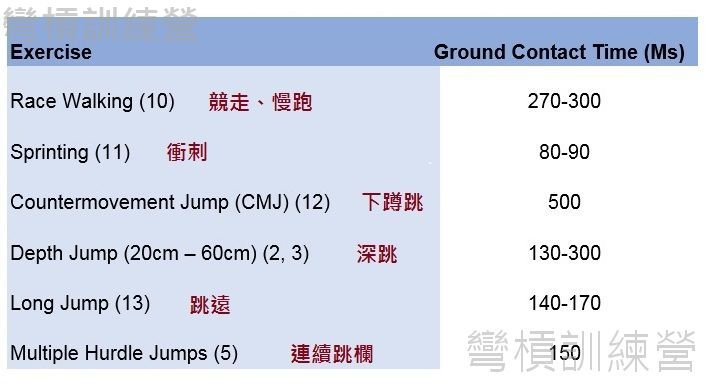
.
依舊以跳躍為例,Kubo et al.的研究指出一般人能在跳躍前靠著反向動作或肌肉從離心到向心所儲存的能量增加跳躍高度,有反向動作的CMJ和DJ較沒有反向動作的SJ增加的幅度可以達到平均3~4cm左右;而同批受測者經過12周的增強式訓練(plyometric training),整體的跳躍能力都有進步,有反向動作的跳躍比SJ還要高出5~7cm的高度 [6]。
從上述內容衍伸出來的意義即是SSC能有效增加力量 與SSC經過鍛鍊後可以更加有效率的應用,但是原因為何? 接著的段落就要來針對SSC的益處做討論,也會點出SSC的原理,並會以上述的三種蹲跳動作和跑步來舉例。
SSC的增益效果與力量-速度曲線*:
*[註2.]:在看這段之前,建議先複習 [肌纖維的科學3: 離心、向心、等長收縮衍伸力量-速度的關係],會比較好理解這一段的內容: https://wordpress.com/view/barbendstrengthcenter.wordpress.com。
在[肌纖維的科學3: 離心、向心、等長收縮衍伸力量-速度的關係],林教練向大家介紹了力量-速度的關係,也詳細的講解了力量-速度曲線(F-V curve),這與本篇文章的關係是,科學家能針對SSC運動做相應的力量測量,就能繪出屬於SSC運動的力量-速度曲線。 如{圖 7.}所示,圖上的小白圈代表每次前臂肌肉做最大努力抵抗不同阻力的反覆測試,經由許多次測試後才能畫出如此漂亮的力量-速度曲線(實線);但是在虛線部分只包含向心運動的測試,此部分的測試是每次向心收縮前先讓前臂受到被動阻力拉長後,才由肌肉主動出力抵抗,每次實驗結果由叉叉表示。最終畫出來的向心曲線比原始的曲線還要向右移,代表著力量與速度都有顯著增強 [7]!
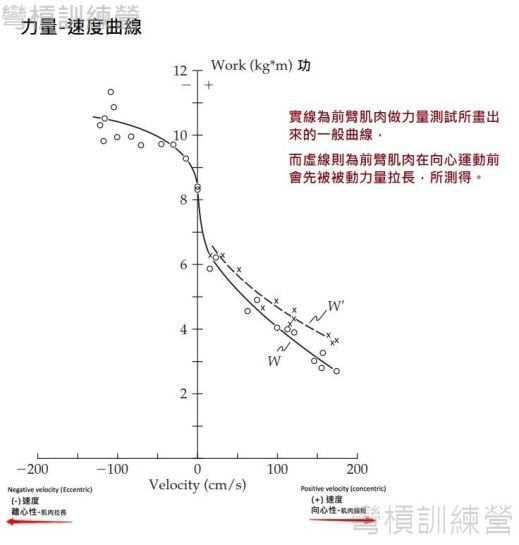
.
傳統的力量-速度曲線是利用分離的肌束做測試而畫出,在人體孤立肌群測試時也能畫出類似傳統的曲線; 但是當科學家想了解一個動作而非單一肌肉的狀態時,就能用瞬時力量-速度曲線(Instantaneous F-V curve)了,此曲線所注意的通常不只是肌肉本身,而是專注在一個"動作中(可以是走或跳)"該肌肉的速度與力量,因此畫出來的曲線會很大程度受到動作本身的影響,所以通常都能看到瞬時力量-速度曲線會被涵蓋在傳統的力量-速度曲線之中,因為人們不常以最大努力來執行一般的動作 [8]。
{圖 8.}的瞬時力量-速度曲線是受測者被要求做連續跳躍(hopping)時所測得,舉例來說 跳繩就是連續跳躍的一種,此實驗的目標組織是小腿後側肌群(triceps surae muscle group)所共用的阿基里斯腱(Achilles tendon),這實驗是利用小腿肌群收縮後所產生的肌腱張力來繪圖。 實驗結果能發現連續跳躍的瞬時力量-速度曲線有一部份超出了傳統力量-速度曲線,意思是連續跳躍可以發揮出比純做最大向心運動時還要大的肌腱張力,所以肌肉在連續跳躍時所產生的張力會比較大,而在比較遠端的髕骨韌帶張力則是包含在小腿肌群的傳統曲線之中,代表股四頭肌在連續跳躍時的出力幅度沒小腿肌群大 [9]。
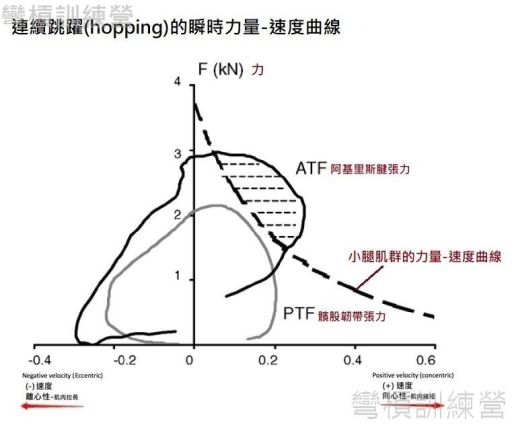
. 上述的結果都證實了有應用到SSC的動作能發揮出更強更快的效果,有這些實驗室的證據,相信大家就不會再有疑惑了。
伸展-收縮循環原理:
大量應用SSC的動作技術在連鎖健身房較為少見,反而是田徑、馬拉松、武術、體操等等力求操作者做出連續動作的運動更加良好的應用此機制 {圖40.}。SSC被證實能提升身體做動作時的能量使用效率,更能透過組織內儲存的能量與牽張反射(stretch reflex)使動作速度更快、力量更大[1、3、6]。 值得注意的是,在跑步的案例中,肌肉在觸地前會出現預活化(pre-activation),{圖9.}中顯示科學家將肌肉的預活化訂為SSC的一個階段,但是此階段只存在於連續動作中(例:跑步、跳繩)或是在做DJ時,當操作者從高處落下,明明身體還沒承受力量也還未跳起,肌肉卻已經被神經活化了,有此可見CMJ等等有應用到SSC的不連續動作中是沒有預活化階段的。
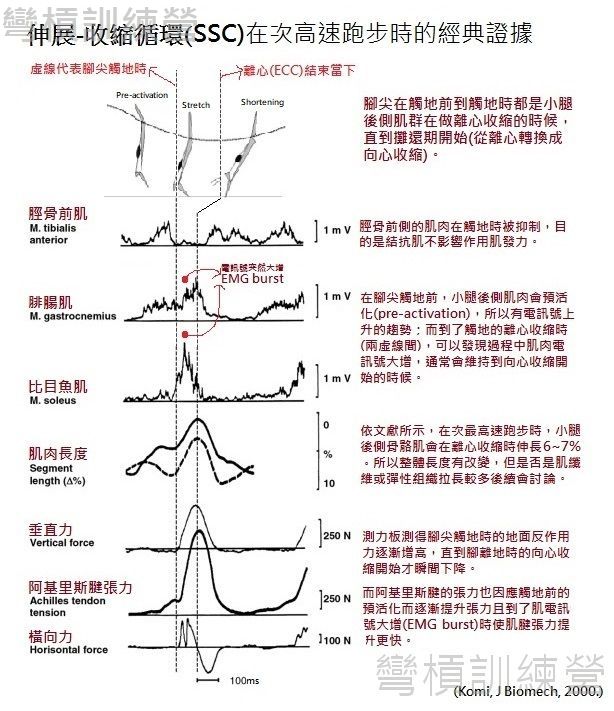
一張圖勝過千言萬語!{圖10.}是出自一篇經典論文的數據,它很全面地以一張圖解釋了次最高速跑步中存在SSC的證據,而這些概念在後續能沿用到所有連續動作之中,此圖涵蓋了大部分SSC的相關機制,包含 1.)肌肉在觸地前即會預活化,可在肌電圖(Electromyography, EMG)的訊號看出端倪; 2.)觸地時的離心動作短且速度快; 3.)攤還期持續時間短,此案例的攤還期約花了100毫秒(ms);額外可以得到的訊息還有 4.)骨骼肌在離心階段確實有長度改變,衍伸出的訊息是彈性組織在SSC扮演的角色; 5.)作用肌在離心階段有出現肌電訊號突然大增(EMG burst)的情況,這是神經反射的增益效果 [9]。
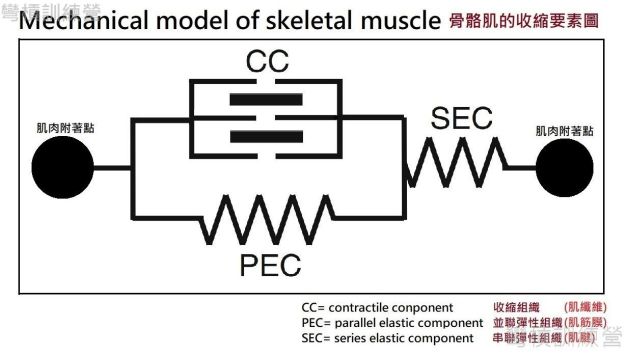
1 *肌腱儲存的彈性位能 (Muscle-tendon elasticity)
理解SSC前的第一步是必須理解骨骼肌的收縮要素{圖11.}。圖中提出骨骼肌主要有三個收縮要素,首要為產生張力的收縮組織(contractile component, CC),而並聯彈性組織(parallel elastic component, PEC)則是與收縮部分平行並緊密連結的部分,第三個收縮要素是串聯彈性組織(series elastic component, SEC),此組織銜接收縮組織和並聯組織 [10]。以此概念圖做類比,收縮組織即為肌纖維,與肌纖維平行排列的並聯組織就是肌筋膜,並連的意義在於肌纖維幾乎都附著在筋膜組織,而筋膜組織最後會匯聚在肌腱處(與成分大致相同的肌腱)做串連,腱組織就是並聯彈性部分。 功能上是由肌肉產生張力,而筋膜和肌腱組織則是抵抗張力;當肌纖維收縮時,周圍無收縮能力的彈性組織會被肌纖維給牽動並拉長*,彈性組織使肌纖維產生的張力能傳導到骨骼上,這是骨骼肌做出動作的方式。
*[註3.]:肌腱與筋膜皆是黏彈性組織(viscoelasticity),能在形變(deformation)時保持彈性並在受力後復原,除非形變到達組織的降伏點(yielding point)才容易斷裂且無法回復原狀。
.
SSC儲存的彈性位能主要在串連和並聯彈性組織之中。與彈性組織相較之下,肌肉並不善於儲存彈性位能,但由於肌肉與肌腱連結在一起,理論上肌腱會承受肌肉主動產生的所有張力,簡單說來就是這些黏彈性組織 誰能被拉到最長就能儲存最多彈性位能。 但這情況還要視「組織剛性」*(tissue stiffness)而定,舉例來說,當我們在做靜態伸展時,由於肌肉並無主動出力,因此肌筋膜的剛性來得比肌腱還要小非常多,因此大部分的形變都發生在以筋膜為首的並聯彈性組織;反之當肌肉出力提升本身剛性時,與肌肉緊密排列的筋膜組織的剛性會超過肌腱組織,因此才能拉動本身剛性較強的肌腱 [11]。 此段的結論是,處於SSC的肌肉在預活化和離心階段會提升本身的肌肉剛性(muscle stiffness),才能牽動肌腱並提升肌腱剛性(tendon stiffness)*,且在離心階段時肌肉-肌腱複合體(muscle-tendon unit)會被拉長,便會儲存彈性位能於其中,到了向心階段便與肌纖維收縮的力量一同釋放出來[6、12]。
一直在2007年以前,對於處在SSC的肌纖維是否有離心-伸長後緊接著向心-縮短存在爭議,原因是前期的文獻並沒有在次最高速跑步中實際觀察到進行SSC運動的肌纖維在離心階段有伸長,相反的是只有等長收縮被觀察到,當時的學界認為在離心階段只有肌腱儲存了很高的彈性位能,這是讓SSC有著增力效果的主因 [9]。 當時是用低頻率(25-50 Hz)的超聲波(ultrasound scanning)進行研究才會產生此誤解,因為科學家沒有在低頻影像內觀察到肌纖維被拉長,只有肌腱組織有被延展開,才有肌纖維從預活化階段到離心階段都在做等長收縮的認知 [13、14]。 但是到了2007年Ishikawa和Komi使用更高頻率(97Hz)的超聲波,觀察到了肌纖維在預活化階段保持等長收縮,但是觸地時的離心階段有短暫的肌肉伸展,且肌纖維在伸展後緊接著測量到牽張反射(stretch reflex)的擴增肌電訊號 [15]!
這小節的結論很簡單,肌腱透過預活化階段和離心階段的肌肉收縮提升肌腱剛性,且此時期也因為肌肉-肌腱複合體被伸展使肌腱儲存了彈性能量,到了向心階段便與肌纖維收縮的力量一同釋放出來。
.
*[註4.]:組織剛性(Δforce/Δlength),以肌腱剛性為例,代表肌腱受力的變量除以長度的變量,值越大代表剛性越強。組織剛性被認為是讓收縮組織產生張力、和使黏彈性組織提供如彈簧般(spring-like)的彈性,剛性高的物體有較良好的傳力效果,而組織剛性隨接觸面等等的改變就是本體感覺應對環境的方式 [16]。
*[註5.]:此文章提到的剛性的英文是stiffness,這與一般民眾苦惱的肌肉僵硬的stiffness是相同的字,但是它們代表的意義大不同! SSC情況下產生的組織剛性在運動結束後就會回復鬆軟有彈性的狀態,不會像肌肉僵硬是生活中無時無刻都因為排列問題而硬梆梆的。
2 *橫橋的活化態* (The active state of cross-bridges)
*[註6.]:在這段開始前,讓我們回憶肌纖維的顯微收縮結構,肌纖維內的蛋白複合體-細肌絲是相對動態的構造,其上方有能讓結構穩固的粗肌絲連結的位置,兩者的蛋白接合位所結合起的構造就是橫橋(cross-bridges),橫橋靠著消耗能量的方式產生抽擊(power stroke)來拉動細肌絲,這就是肌纖維收縮的方式。
詳見[肌纖維的科學1: 肌肉收縮]: https://wordpress.com/view/barbendstrengthcenter.wordpress.com。
橫橋的活化態是肌纖維正在產生張力抵抗外力的狀態,詳細的意思是: 當粗肌絲上的肌動蛋白(actin)被粗肌絲的肌球蛋白頭(myosin head)給牢牢接上形成橫橋,且這些橫橋正在抵抗外在力量,更廣義的活化態還包括再向心階段前給予肌肉產生力量的一段時間(在SSC的運動中是離心階段和攤還期) [17]。
活化態剛好是肌肉進行等長收縮和離心收縮會產生的狀況。 特別是在CMJ等等有反向動作的跳躍可以有較佳的活化態,因為反向動作就是要作用肌做離心收縮以抵抗體重造成的重心下沉,因此肌肉會在反向動作前和動作中搭起橫橋,這一部分的橫橋能保留到向心階段,與向心階段時快速新建的橫橋一起產生力量; 與CMJ相反,SJ因為沒有反向動作,操作者只能在已經下蹲一段時間(等長收縮)後用力跳起,因此操作者的橫橋只能在此向心階段搭起。 或許我們能簡單地想成,反向動作因為是抵抗重力,所以要生成較多的橫橋,而SJ初期的等長收縮因為只需維持姿勢,所以只要少量橫橋就能維持,所以保留到向心階段的橫橋量本身就有差異, 假設兩種不同的跳躍的在向心階段出相同的力,那搭配上向心階段時生成的新橫橋,就會產生很大的力量差異。 這概念還能套用在DJ這種從高處下躍並用力跳起的類型,它因為從高處落下,身體在離心階段需要抵抗的重力還比CMJ高,所以能跳得更高[17、18]。
橫橋的活化態是比較不受科學家重視的一塊,但他實際上也是功不可沒。
(我個人覺得不受重視的原因是因為這只能用電腦模型來模擬,所以沒有實際的實驗成果,且我提到的是較狹義的部分,更廣義的是我看不懂的天書,又或許我對整個活化態的認知其實都不完整,所以大家若發現”活化態”這部分有錯,請別吝嗇你的發現,一定要告訴我!)*
.
*[註7.]:活化態這機制在科學文獻以外是幾乎看不到的,網路上唯一有提及的就是美國知名實務訓練家Owen Walker,他在他的文章中還提到,要優化活化態的方式是在限度內進行較久的離心階段和攤還期,因為他認為這樣可以使更多的橫橋生成,但是大家在實際操作時千萬不要把時間拉太長,不然有可能會本末倒置,因為我們所儲存的能量會以熱量形式散發掉。
Stretch-Shortening Cycle: https://www.scienceforsport.com/stretch-shortening-cycle/#toggle-id-1。
3 *神經的調控 (Neurophysiological regulation)
緊接著來討論神經要素,我們在前段有提到離心階段時的肌肉快速伸展能讓肌電訊號產生增幅,這被廣泛認為是牽張反射所提供的增益效果(potentiation),可參考{圖10.}。
產生牽張反射是由「肌梭(muscle spindles)」跟著肌纖維一起被離心-伸長所產生{詳見附錄1.},傳統上認為牽張反射的產生可以增加運動單位(motor units)的徵招,使得SSC最後一段的向心階段能有更多肌纖維的參與,但這目前是有爭議的。 因為目前的科學研究還沒有取得最直接的證據,相關的研究都是肌電圖實驗,通常是讓受測者接上表面電極後開始做運動。 測量牽張反射是否存在於運動中 對科學家很簡單,只要在運動中發現肌肉的訊號有特殊的上升情形就好,最好是來個抑制牽張反射的控制組,這樣更有說服力,因此科學家就以此設計實驗得到了如他們預期的結果。 首先是科學家用踏板針對小腿肌肉做被動伸展測試,結果是快速的透過踏板被動地伸展小腿,導致阿基里斯腱的張力較原本增加了數倍,其中也發現EMG訊號在伸展中期有增幅 更伴隨肌腱張力急遽上升的現象,而慢速伸展則完全無反應 {圖 12.} [19],因此一般的被動伸展是足以引起牽張反射的。這研究對SSC運動的意義不太大,但也是一個基石,畢竟此研究使用的快速伸展是以每秒69度再轉動關節,這速度在SSC運動中是可以達到的。
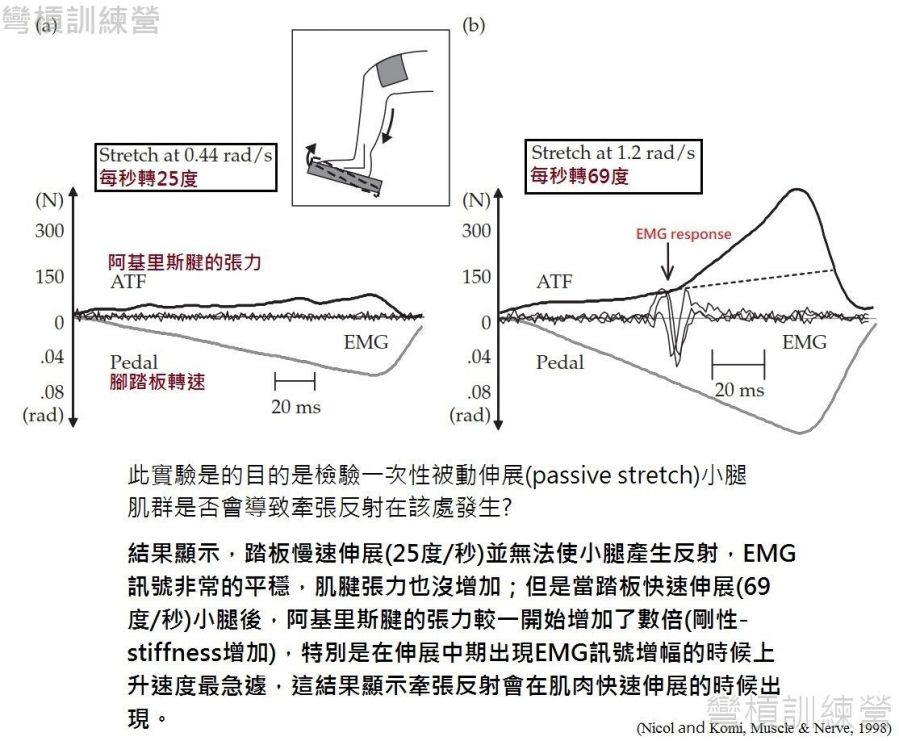
.
接著是真正有應用到SSC的運動測試,受測者被要求在測力板(force plate)上做深跳(DJ from 60cm height),在跳躍時會一併測量肌電數據。受測者表現出的形式與前述的SSC特徵一致,從60cm跳箱下躍時,肌肉有預活化情形;且當受測者在觸地到跳起來的離心階段更出現肌電訊號的突然增幅,這些肌群產生肌電訊號增幅的時段緊接著向心階段{圖 13.} [7]。 做出類似結果的實驗也多如繁星[7、13、14、15]。
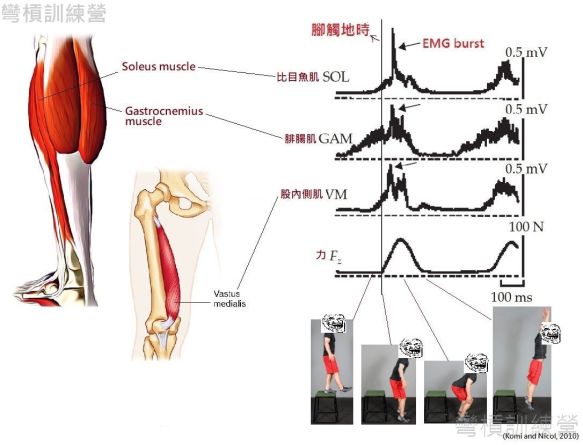
.
Dietz等人在1979年進行了比較全面的研究,他讓受測者在跑步前先行測量腓腸肌的最大等長收縮之肌電訊號,發現腓腸肌在跑步時的肌電訊號比最大等長收縮時的還要高2~3倍{圖 14 a.},此受測者的跑速皆設定在最高速的70%。 之後,科學家又對受測者進行局部缺血(ischemia)的處理,使腓腸肌肌梭內的Ia感覺神經短暫喪失功能後,再度執行腓腸肌最大等長收縮及跑步測試,這個實驗結果相當讓人驚訝,腓腸肌在缺血處理後的跑步肌電訊號與最大等長收縮的訊號沒有明顯的差異,但是卻與缺血處理前的肌電訊號相差2~3倍{圖 14 b.};科學家在論文中指名,肌電訊號的增幅是發生在腳尖觸地之後的35-45微秒之間,這是相關研究中常提及的反射時間,因此搭配上實驗組與對照組之間的比較後,發現牽張反射確實是存在跑步之中,也更進一步能推論是存在SSC運動中[7、20]。 但是,此研究的受測者儘管在肌電訊號上有顯著差異,但是在跑速沒差異(理論上是出力相同)的狀態下,且腓腸肌在最大等長收縮時與在跑步時的性質相差極大,因此此研究並無法明確告知腓腸肌產生的力量是否有不同。
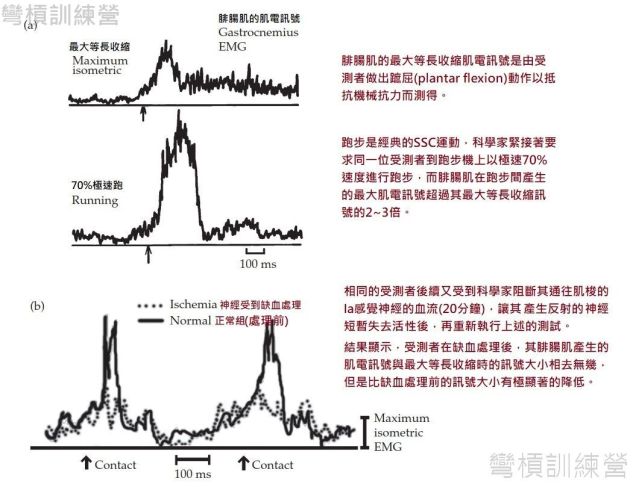
.
雖然目前的研究明確指出SSC會伴隨著牽張反射一同發生,但是他們實際上的連結還有待進一步釐清。 主因是目前無法測到單一牽張反射的訊號,雖然有肌電訊號的增幅,但是增幅的訊號是與肌肉收縮的肌電訊號混雜在一起;而且隨著肌電訊號增幅的外部效果也無法準確測得,因為向心收縮還包含著自主性;其他影響因素還包括參與SSC的組織特性會影響力量的增益,所以目前依舊是處在相對模糊的狀態[11]。
儘管目前對於SSC運動是否受益於牽張反射所提供的增力效果還相對模糊,但是現有的文獻皆一致性地指出有無牽張反射的發生會與肌腱剛性有直接相關[7、9]。以其中一篇經典文獻為例,此研究是用腦部切除的貓做神經肌肉分離手術與否,讓控制組的比目魚肌(soleus muscle)保有反射能力而實驗組的貓則是沒有反射能力;實驗以測力裝置對比目魚肌施加操作力(operating force)的同時給予電刺激來維持等長收縮,過程中會逐漸增加操作力,科學家以此觀察肌腱剛性。 結果發現在相同單位的操作力下,保有正常神經反射的肌腱剛性比缺乏反射能力的還要高出許多,且趨勢是以初始的較低操作力就能產生不小的剛性提升,而到較高的操作力雖沒造成剛性以線性地上升,可是依舊展現出提升的態勢直到抑制反射出現; 這是無反射組所沒有的反應,且無反射組的肌腱剛性在肌肉強直(tetanus)之前都是以線性趨勢提昇{圖 15.} [7、21]。
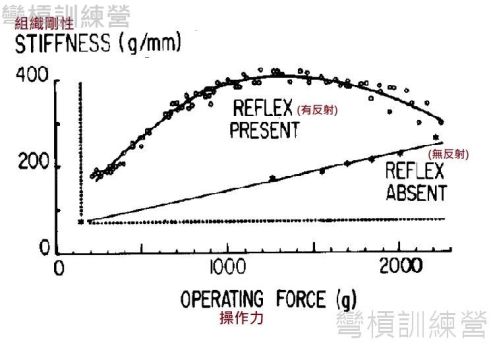
.
進行SSC運動時,有牽張反射發生(不管是被動還主動)的組織能發現剛性會大幅提升,對於身體面對環境有著正向幫助,這是相當合理的身體機制,若運動時身體沒產生足夠的剛性,對於觸地的應力將完全會由第一線的骨骼及重要的結構軟組織吸收,因此牽張反射的介入可以防止未在預活化時增加足夠剛性的肌肉做出退讓(yeilding) [22]。 離心階段中發現肌電訊號上升伴隨著組織剛性快速提升,而是否牽張反射的啟動能刺激更多肌纖維加入向心收縮的行列還無法完全確定,雖然組織剛性的提升能很大的代表肌肉是處在收縮狀態,因此依照前人的研究結果,我們可以合理地認為牽張反射在SSC的運動中有提供增益效果。
總結:
在運動中甚至是日常生活中,SSC是無所不在的,它能讓運動者發揮出比單純做向心收縮還要力量更強、速度更快的效果,而SSC不只能增進爆發力運動的表現,也能對耐力運動有幫助。完整應用到SSC機制的運動是以快速的伸展-收縮運動才有(有些動作還不一定會是完整的SSC運動),而SSC運動對未經訓練的人來說相當危險,但是對有肌力訓練基礎的人來說是進步的利器!
上述組織彈性和神經調節機制是被大家廣泛認同的SSC機制,而活化態是比較少人提及的另一個可能性,相信大家看過之後能對SSC有更深入的理解,另外由於篇幅有限,本文章就不列出其他可能參與的機制,其他機制的重要性是有,但是都沒我在文章中所提及的這麼有力。
SSC對骨骼肌的增益效果來自於1.)預活化階段的骨骼肌剛性提升和2.)離心階段時的牽張反射更讓肌腱剛性大幅提升;且此時期也因為3.)肌肉-肌腱複合體被伸展因此讓肌腱儲存了彈性能量;而在整個4.)離心階段也讓大量的橫橋能充分搭起,這上述的機制幫助運動員在最終的向心階段能快速地釋放出更強的力量。
.
最後,林教練列舉一些SSC出現在運動的例子給大家參考:
*短跑(sprint) {圖 16.}– 短跑是沒有反向動作的SSC運動,因為它是連續動作。 我們來看看它應用到的部分,在衝刺中的每次髖屈曲,都是為了能讓髖伸展所做的準備;而每次膝關節伸展都是在為下一次膝關節屈曲做準備,因為跑步時肌肉的每次牽張-縮短會儲存了大量的彈性位能,才能完成此複雜的動作。
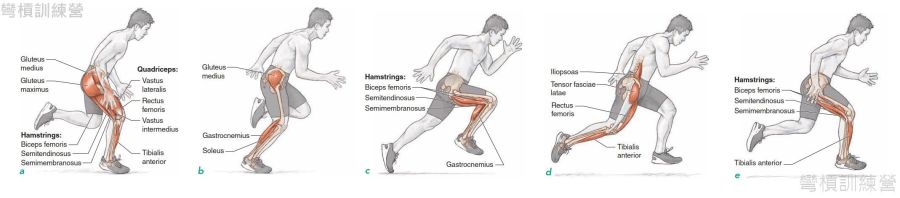
.
*分腿上挺(Split jerk)- 舉重是會大量應用SSC的技術性運動,在上挺之前,身體會做出下蹲的反向運動,迅速讓重心下降約自己身體的十分之一的高度後,馬上用力瞬發使重心抬升,一併將槓鈴給推至頭頂。
.
*懸垂抓舉(Hang snatch)- 舉重是會大量應用SSC的技術性運動,它那複雜的動作能分解成數種專項輔助動作來幫助訓練,這懸垂抓舉就是一例,當操作者將槓鈴給抓到腰際高度時,馬上做出幫助身體積蓄彈性位能的反向運動,並配合抓舉的動作技術,一次性地將槓鈴給帶到頭頂。
.
*射門(Kicking a soccer)- 足球一直是風靡全世界的團隊性運動,大家最愛看的就是射門得分的那一刻。 足球員在將球踢出去之前,可以發現它會將整條腿給向後伸,這就是踢球前的反向動作,此舉為大腿伸肌積蓄了彈性能,最後隨著向心階段一起將球給踢出去。
附錄:
1. 牽張反射
牽張反射最著名的一例就是 膝跳反射(knee-jerk reflex)*,當膝關節屈曲且輕鬆懸垂於桌緣時,拿一個尖頭槌敲擊髕骨韌帶(patellar ligament),就能使身體產生反射,這時會看到被敲的那條腿用力的往前踢直。這機制相當簡單,我們的骨骼肌內含相當多的本體感覺受器(proprioception receptor),導致膝跳反射的受器就是肌梭(muscle spindle)。 由於尖槌敲擊膝蓋韌帶,造成腱組織凹陷並拉扯周圍組織,此舉造成股四頭肌內的肌梭被拉扯,最後啟動反射路徑。 反射路徑初始是由肌梭的Ia感覺神經(Ia sensory fibers)連結到脊髓的α運動神經元(alpha motor neurons),然後激發股四頭肌急速收縮;另外Ia傳入神經也會連結到脊髓中的抑制神經元(inhibitory neurons),此抑制神經會讓腿後腱肌群(hamstring)保持在鬆軟的狀態,以利股四頭肌順利出力{圖17.} [23]。
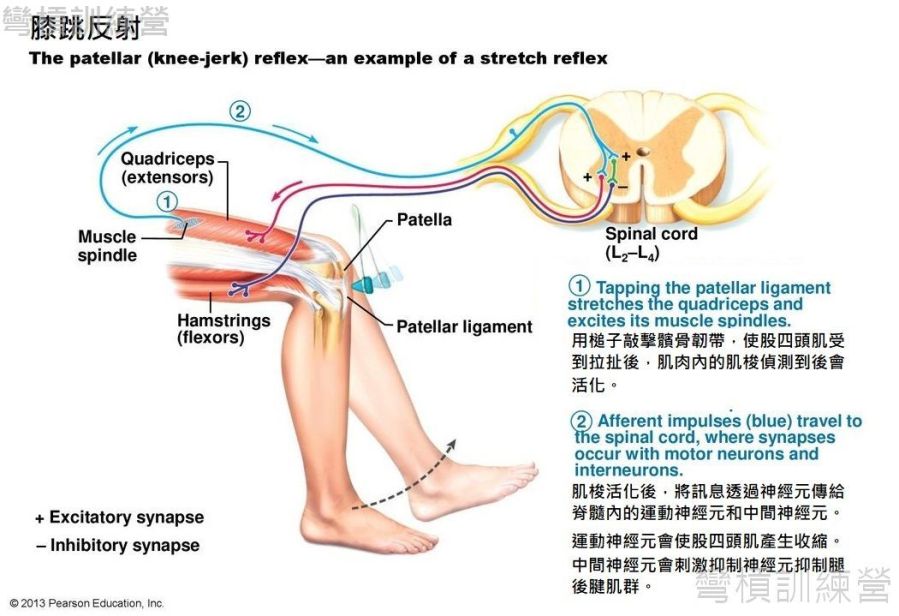
.
肌梭是散布在骨骼肌內的本體感覺受器之一{圖18.},形狀類似紡錘或繭,兩端連接在肌纖維、筋膜及肌腱上,因此能感受到骨骼肌的牽拉。 我們能自主控制的肌纖維又叫做梭外纖維(extrafusal fibers),而肌梭內的梭內纖維(intrafusal fibers)又有兩種類型,分別為核袋纖維(nuclear bag fiber)和核鏈纖維(nuclear chain fiber),每個肌梭內約有2個核袋纖維和4~5個核鏈纖維。這些肌纖維有比梭外纖維還要密集的神經連結,例如偵測動態和靜態時的骨骼肌長度的神經是Ia感覺神經(主要纏繞在兩種梭內纖維的中央),而另一種是Type 2感覺神經纖維(type II sensory fiber),能偵測肌梭受到的張力並提供本體感覺的輸出[24]。
這些內容也相當有趣,未來可以多做介紹。
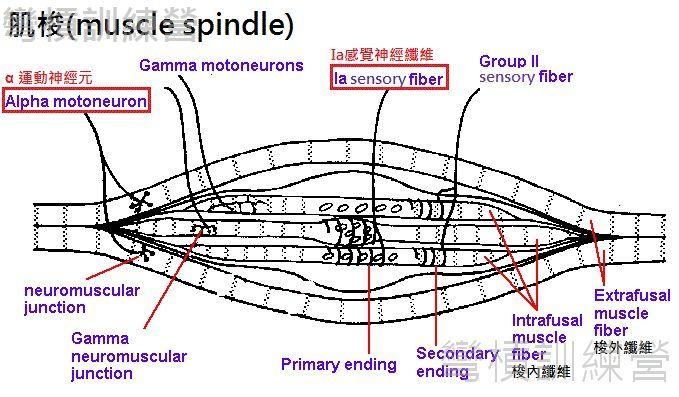
*[註8.]:膝跳反射是比較基本的單突觸反射(monosynaptic reflex),意思是最簡單的反射,它的神經傳導並沒有經過其它的中間神經元(interneurons);而反射機制是相當複雜的,相似的反射也有更高層級的多突觸反射(polysynaptic reflex)有些有肌梭參與有些是不同的受器所引發的,這些有趣的功能可以在參考資料[25]做大略的閱讀。
..
2. 梭內纖維會參與牽張反射提供給骨骼肌的增益性嗎?
林教練好久以前還一度以為牽張反射所帶來的增益效果是由梭內纖維與梭外纖維的一同收縮所帶來的!? 當然這在數十年前就被證明是錯的,而且肌梭的功能也早已被闡明並寫進教科書內,梭內纖維不會與梭外纖維共同出力,所以接下來的討論也只是因有趣而為之,非以事實蓋之。
在討論梭內纖維能出多少力之前,要先了解其密度,但是難題又來了,不同部位不同肌肉所含有的肌梭數量又完全不一致,目前現有的資料如{圖19.}中列出部分肌肉的平均每克肌肉所含的肌梭數量,而肌梭對蛋白質量更高的肌纖維來說是輕如鴻毛[26],所以這些文獻都不會去管每克肌肉所含的肌梭重量。那現在假設一塊肌肉重一公斤,以{圖19.}提供的最高肌梭密度來看,這塊肌肉所含的肌梭數量也不超過17500個,這些輕如鴻毛的肌梭與梭外纖維的質量相比是微不足道的,但似乎很難以此方法界定肌梭能否出很多力。
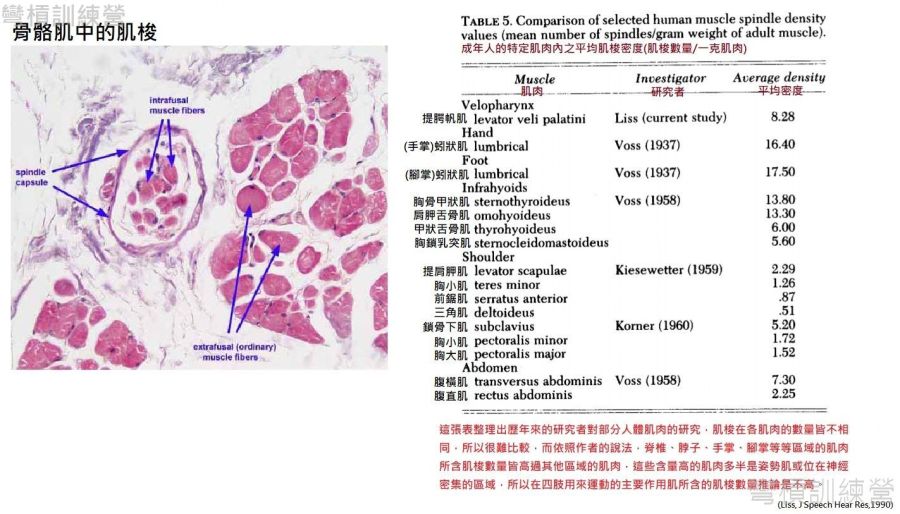
.
接著就讓我懶惰一下,因為相關文獻已經找不到了,直接翻譯由知名教授Mann MD所撰寫的網路教科書的部分段落 [24]:
肌肉中所有的梭內纖維一齊收縮並不會產生可測量到的力,主因是梭內纖維比梭外纖維還要少上許多,且梭內纖維在結構上比起梭外纖維還要短且小。梭內纖維的長度約為4-7毫米(mm),而梭外纖維以整體骨骼肌層面來看約為3到50公分不等,整個宏觀視野下所看見的肌纖維全都是屬於梭外纖維,且梭外纖維在收縮時最多能減少40%的原始長度,此收縮長度若套用在梭內纖維中(7mm->4.2mm),其整體收縮長度也無法跟巨量的梭外纖維相比較;另外,單一梭外肌纖維直徑約為60微米(um),而梭內肌纖維則是10微米(um),以骨骼肌直徑與產生力量成正比的概念看來,梭外肌纖維能產生的力量是遠遠超過梭內肌纖維能生成的力量,以上概念整合後,就不難發現為何梭內纖維並不會產生多少力量了。
.
3. 預活化是前饋(feedforward)而反射是回饋(feedback)
此文章所提到的反射是身體面對環境所產生的回饋,回饋控制(又稱為reactive control)整合了周邊反饋的信息並及時幫助身體調整當下動作與加固,這邊提到的周邊反饋可以是周邊神經的感覺輸入,也能是反射,總之回饋是一個應對當下的機制[27]。而前饋則是相反的意思,前饋控制代表人體在動作前所做的計畫或是預測,因此並無包含周邊系統所提供的即時調整[27],前饋在本篇文章中的最佳例子為肌肉預活化{圖9.}。
前饋控制是經由不斷的練習所產生,例如: 讓受測者在軟地上不斷跳躍,此時大腦會透過回饋機制學習到新的內部模式(internal model),所以後續要求受測者在不同平面上執行相同工作,就很有可能在運動初期產生錯誤(error),但是不久之後又能產生正常反應。 這例子表示動作輸出必須被整合與校準,並以此適應多變的環境。 因此,中樞神經系統理論上會記錄四肢及身體動態的內部模式,這些模式能作為中樞神經系統面對類似情境的板模,讓受測者能針對類似情境做出相應的動作輸出;但是當受測者被要求在不同的情境,讓身體面對不同的生物力學特性下,再度執行類似動作,就有可能產生動作失誤,因此內部模式是需要更新的[22],這邊的更新用白話文來說就是「練習」和「學習」的不斷循環!
前饋在有落地的高速運動中特別重要,例如深跳(DJ)這種需要肌肉預活化的動作。科學家有提出如何發現前饋的存在,可以要求受測者在正常情況下做DJ後,接著矇眼做DJ,起初的動作模式相當怪異,但是在幾次嘗試後就能做出與睜眼時一致的動作,但是當研究者突然改變跳躍高度來蒙混受測者,偷偷地提高從60公分深到20公分深,受測者卻連一個完整的跳躍都做不到 [22],這有點像不認真走樓梯的情況,眼睛沒預估情勢,明明已經到地面了,身體還繼續往下走,最後就是突然來個踉蹌作結。
{圖20.}是附錄3的總結,其實SSC就是前饋與回饋的整合運作。
這系列到第五篇了,每篇雖然都有獨立的內容,但是沒稍微看過前幾篇的新讀者或許會越來越看不懂呢; 就連作者我自己都時常有這感受,畢竟要把這些知識整理得有連貫性,必須花費不少功夫,因為我必須在規劃文章內容前先了解整體的知識,包括生理學、運動生理學、解剖學,是說林教練本身是解剖學研究所出生,還好大部分的知識都看過,整理起來相對方便。 就希望一路發樓的讀者們能在我的訓練營內一路行進,在這訓練營當中能理解練身體到底在練什麼,這將是跳脫練給鏡子看的第一步。
參考資料:
- Nicol C, Avela J, Komi PV. (2006). The stretch-shortening cycle : a model to study naturally occurring neuromuscular fatigue. Sports Med, 36(11), 977-999.
- Komi PV, Bosco C. (1978). Utilization of stored elastic energy in leg extensor muscles by men and women. Med Sci Sports, 1978, 10(4), 261-265.
- Gehri D J, Ricard M D, Kleiner D M et al. (1998). A comparison of plyometric training techniques for improving vertical jump ability and energy production. J Strength Cond Res, 1285–1289.
- Cavagna GA, Heglund NC, Harry JD, Montovani M. (1994). Storage and release of mechanical energy by contracting frog muscle fiber. The Journal of Physiology 481, 689–708.
- Walker O. (2016). Incremental DJ-RSI test. {圖6.}
https://www.scienceforsport.com/incremental-dj-rsi-test/ - Kubo K,Morimoto M,Komuro T,Yata H,Tsunoda N,Kanehisa H, Fukunaga T.(2007). Effects of Plyometric and Weight Training on Muscle-Tendon Complex and Jump Performance. Medicine & Science in Sports & Exercise, 39(10), 1801–1810.
- Ishikawa M and Komi PV. (2010). Neuromuscular Aspects of Sport Performance, Volume XVII. 10.1002/9781444324822.ch2. {圖 7.} {圖 13.} {圖 14.}
- Sonnenblick EH. (1965). Instantaneous Force-Velocity-Length Determinants in the Contraction of Heart Muscle. Circ Res, 16, 441-451.
- Komi PV. (2000). Stretch-shortening cycle: a powerful model to study normal and fatigued muscle. J Biomech, 33(10), 1197-1206. {圖 8.} {圖 9.} {圖 10.}
- Siebert T, Rode C, Herzog W, Till O, Blickhan R. (2007). Nonlinearities make a difference: comparison of two common Hill-type models with real muscle. Biol Cybern, 98(2),133-143. {圖 11.}
- Turner AN, Jeffreys I. (2010). The Stretch-Shortening Cycle: Proposed Mechanisms and Methods for Enhancement. Strength and conditioning journal, 32(4), 87-99.
- Kubo K, Kawakami Y, and Fukunaga T. (1999). Influence of elastic properties of tendon structures on jump performance in humans. J Appl Physiol, 87, 2090–2096.
- Fukunaga T, Kubo K, Kawakami Y, Fukashiro S, Kanehisa H, Maganaris CN. (2001). In vivo behaviour of human muscle tendon during walking. Proc Biol Sci, 268(1464), 229-233.
- Ishikawa M, Finni T, Komi PV. (2003). Behaviour of vastus lateralis muscle-tendon during high intensity SSC exercises in vivo. Acta Physiol Scand, 178(3), 205-213.
- Ishikawa M, Komi PV. (2007). The role of the stretch reflex in the gastrocnemius muscle during human locomotion at various speeds. J Appl Physiol (1985), 103(3), 1030-1036.
- Houk JC. (1979). Regulation of stiffness by skeletomotor reflexes. Annu Rev Physiol, 41, 99-114.
- Bobbert MF, Casius LJ. (2005). Is the effect of a countermovement on jump height due to active state development? Med Sci Sports Exerc, 37(3), 440-446.
- Van Ingen Schenau GJ, Bobbert MF, de Haan A. (1997). Mechanics and Energetics of the Stretch-Shortening Cycle: A Stimulating Discussion. Journal of Applied Biomechanics, 13, 484-496.
- Nicol C, Komi PV. (1998). Significance of passively induced stretch reflexes on Achilles tendon force enhancement. Muscle & Nerve, 21, 1546–1548. {圖 12.}
- Dietz V, Schmidtbleicher D, Noth J. (1979). Neuronal mechanisms of human locomotion. J Neurophysiol, 42(5), 1212-1222.
- Hoffer JA, Andreassen S. (1981). Regulation of soleus muscle stiffness in premammillary cats: intrinsic and reflex components. J Neurophysiol, 45(2), 267-285. {圖 15.}
- Taube W, Leukel C, Gollhofer A. (2012). How neurons make us jump: the neural control of stretch-shortening cycle movements. Exerc Sport Sci Rev, 40(2), 106-115. {圖 20.}
- Learn more about Stretch Reflex.
https://www.sciencedirect.com/topics/neuroscience/stretch-reflex - Mann MD. (2011). Muscle Receptors. {圖 18.}
http://michaeldmann.net/mann11.html - Learn more about Polysynaptic Reflex.
https://www.sciencedirect.com/topics/neuroscience/polysynaptic-reflex - Liss JM. (1990). Muscle spindles in the human levator veli palatini and palatoglossus muscles. J Speech Hear Res, 33(4), 736-746. {圖 19.}
- Bastian AJ. (2006). Learning to predict the future: the cerebellum adapts feedforward movement control. Curr Opin Neurobiol, 16, 645-649.
這邊是「彎槓訓練營」,我的目標是以平易近人的口吻將科學知識傳達給大眾,我會對複雜的科學內容做出省略,卻不會刪剪掉或破壞整體脈絡,期待能讓各位讀者從文章的內容學到終身受用的知識。
還請大家多多支持,有問題歡迎留言提問!
粉絲頁連結:(若喜歡這邊,請不要吝嗇你的讚)
https://m.facebook.com/weightliftinginbbsc/
